Происхождение жизни. От РНК‑мира к белкам
03.05.2022
Источник: Троицкий вариант, 03.05.2022, Михаил Гельфанд; Борис Штерн
Тем, кто посетил
сей мир в его минуты роковые, очень важно не забывать о вечных вопросах
— они задают систему ценностей, систему координат, с которой всё происходящее
видно как бы с высоты. Это помогает сохранить трезвый рассудок, твердость
духа и осмысленность действий. Среди главных вопросов такого типа — место
человека во Вселенной: в частности, что такое жизнь, как она произошла,
насколько это уникальное явление.
Три года назад на
страницах «Троицкого варианта» шло обсуждение проблемы возникновения
жизни. Сейчас, по нашему мнению, пришло время вновь поднять эту тему на более
глубоком уровне: она так или иначе связана с вопросом о ценности жизни — вопросом,
который сейчас находится в глубокой тени. Теперь мы возвращаемся к этой
дискуссии в несколько другом жанре — в виде серии интервью со специалистами,
которым есть что сказать и которые умеют говорить внятно.
Это не последние
интервью на тему происхождения и развития жизни и не последняя тема
в рамках вопроса о месте человека во Вселенной.

Борис Штерн
Сейчас будет редкое зрелище. Главный редактор (Борис Штерн) будет пытать своего зама (Михаила Гельфанда), причем главный
редактор — в качестве студента, зам — в качестве профессора. Мы
начинаем довольно большую серию интервью и статей с довольно простым
месседжем: когда плохо вокруг тебя и тебе самому, иногда очень полезно
вернуться к основам и освежить систему координат. Поэтому мы будем
говорить о таких фундаментальных вещах, как происхождение жизни, ее
ценность, Вселенная, экзопланеты, возможность колонизовать другие миры и так
далее.

Михаил Гельфанд
— Я понимаю начало происхождения жизни так.
Появились какие-то структуры, которые умеют сами себя воспроизводить, но
не точно, а с некоторыми отклонениями,
и благоприятные отклонения запоминаются. Это и есть эволюция по
Дарвину. Старт эволюции — начало жизни.
— Эти структуры
в просторечии называют «репликаторы».
— По современным воззрениям, что такое первый
репликатор и как он возник?
— Разных
современных воззрений много. Кажется разумным, что первыми репликаторами были
молекулы РНК. Первые этапы возникновения жизни целиком, конечно,
в пробирке не реализованы, потому что времена не те, но по частям —
реализованы. Сделаны молекулы РНК, которые способны к матричному синтезу,
то есть к тому, чтобы скопировать другую молекулу РНК — это означает, что
им можно подсунуть в качестве матрицы их самих.
— ДНК не способны себя копировать?
— ДНК не может
вообще выполнять никаких каталитических функций. ДНК — штука довольно тупая.
А РНК… Есть рибозимы, за открытие которых Том Чех в свое время
получил Нобелевскую премию: это молекулы РНК, которые являются ферментами. Одна
из возможных для них ферментативных активностей — это матричный синтез РНК. Соответственно,
РНК-фермент читает РНК-матрицу и делает РНК-копию. Там есть тонкие фокусы
с комплементарностью, но бог с ними. Принципиальная возможность существования
РНК-репликаторов показана в пробирке.
Дальше: вроде бы
смоделирована ситуация, когда из раствора того состава, который реалистичен для
ранней Земли, и в условиях, которые реалистичны для ранней Земли,
самопроизвольно возникают достаточно длинные молекулы РНК.
Однако имеется
некоторый зазор. Мы пытаемся перепрыгнуть пропасть в несколько прыжков,
и нам не хватает нескольких промежуточных уступов. Есть две проблемы.
Первая проблема: те РНК, которые получаются самопроизвольно, более короткие,
чем те, которые нужны для матричного синтеза. Вторая: матричный синтез такими
рибозимами неточный. Да, с одной стороны, нам нужно, чтобы были изменения,
чтобы они фиксировались, чтобы было над чем работать дарвиновскому отбору.
С другой стороны — если ошибок слишком много, то у нас вся
наследственность просто растворяется. В предельном случае, если копирование
происходит со стопроцентными ошибками, у нас нет никакого матричного синтеза,
потому что не сохраняется память.
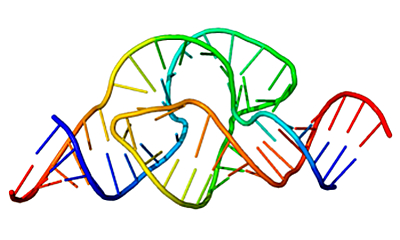
Структура рибозима
— молекулы РНК, выполняющей функцию катализа. «Википедия»
Есть красивое
решение, которое тоже экспериментально реализовано. Оно состоит
в следующем: нам не обязательно, чтобы рибозим был одной-единственной
молекулой; мы можем собрать рибозим из более коротких частей. Так решаются обе
проблемы. Во-первых, если он состоит из коротких фрагментов, мы можем их
синтезировать по отдельности. Они потом сами соберутся за счет комплементарности
между цепями. Спирали РНК, за счет которых образуется пространственная структура,
не обязаны образовываться в одной молекуле, они, вообще говоря, могут
образоваться между разными молекулами. Во-вторых, решается катастрофа ошибок,
потому что ошибки будут распределены по Пуассону, и если участки относительно
короткие, то будут такие фрагменты, в которых ошибок нет или мало,
а дальше нам рано или поздно повезет, и время от времени из них будут
собираться такие рибозимы, в которых почти нет ошибок. Дальше мы всё это
умножаем на колоссальные времена и колоссальное количество попыток
и получаем относительно правдоподобный сценарий возникновения первых
репликаторов.
— Количественный вопрос. У Кунина была
оценка возникновения полного репликатора, и получилось, что вероятность
около 10–1000, причем всё равно, на что нормировать. Нормируем мы на
один акт или на целую планету, там разница в десятки порядков всего,
а не в тысячи. Как я понял, если этот процесс раздробить на
несколько разных независимо работающих рибозимов…
— Нет. Рибозим
один, но он состоит из нескольких частей, которые независимо синтезируются.
— Понятно. Тогда там оценки вероятности сильно плывут?
— Понятия не имею.
У меня ощущение, что эти прикидки настолько отфонарные, что пусть ими
физики занимаются, а не биологи. Я не понимаю, откуда берутся эти
оценки. Вероятность создания самого первого репликатора из случайных наборов
я оценивать не умею. Подозреваю, что никто не умеет, потому что непонятно,
какие исходные данные брать, что на что умножать надо, чтобы получить эту
оценку.
— У Кунина всё просто…
— Кунин много
общается с физиками, его это испортило. И вы даже знаете этих физиков.
— Хорошо. Тем не менее оценка прямая
и лобовая: имеем молекулу такой-то длины…
— Не канает по
тривиальной причине: мы не знаем количество потенциальных молекул, которые были
бы хороши. Вы можете оценить одну конкретную молекулу, но вы не знаете, какое
количество молекул годится. Там несколько порядков тоже можно упустить.
— То есть нужно что-то радикальное?
— Еще раз: этого
никто не знает. Я вижу принципиальную возможность, но я не умею
оценивать времена. И, видимо, никто не умеет. Если у нас получается
10–1000, то всё остальное значения не имеет, потому что вы можете всю Вселенную
напихать молекулами — и всё равно ничего не произойдет.
— Вселенная очень большая, очень.
—
101000 молекул там поместится?
— Поместится, не забывайте про
мультиверс — вселенных бесконечно много.
— А на Земле?
— Нет, конечно. Там 1050, даже меньше, если речь
о молекулах.
— Ну вот. Следует
ли из этих оценок, что есть еще какой-то принципиальный шаг, которого мы не
понимаем, я не знаю. Может быть, да; может быть, нет. Я не умею обращаться
с такими числами. Как вы говорите, Вселенная большая. Дальше начинается всякая
философия в виде антропного принципа, и мы можем заниматься не
биологией, а философией сколько угодно.
— Я считаю, это плохой
стиль — списывать всё на антропный принцип.
— Опять-таки: мы
с Евгением Викторовичем [Куниным. — Ред.] об этом дискутировали.
Списывать на антропный принцип — это, по-моему, интеллектуальная капитуляция.
— Именно это мы писали про энергию вакуума
с Валерием Рубаковым… Едем дальше. Появился репликатор.
Допустим, РНК. Но первая жизнь, которую мы видим, — это клетка. Путь
от РНК до клетки простой или не очень?
— Первая жизнь,
которую мы видим, — это все-таки не клетка, а химический состав
каких-то осадков, который трудно себе представить путем неорганического
синтеза. Никаких клеток мы при этом не видим, насколько я понимаю. Вообще,
я не специалист по этим вопросам.
По существующим
моделям, репликаторы живут в каких-то глинистых породах, где имеются микрокомпартменты
— попросту говоря, поры — вследствие геологической структуры породы, и это
хорошо тем, что появляется понятие коллектива репликаторов, потому что они
ограничены в пространстве и могут взаимодействовать друг
с другом. Это довольно существенная вещь, потому что появляется разделение
функций. Кто-то лучше делает одно, кто-то лучше делает другое, а из-за
того, что они пространственно ограничены, они не растекаются, и функции,
завязанные друг на друга, могут эволюционировать параллельно.
И дальше есть
два принципиальных момента. Насколько я знаю, мы даже не очень понимаем,
в каком порядке они следовали друг за другом хронологически. Во-первых, появление
мембраны. Она улучшает компартментализацию, делает ее более надежной
и дает возможность коллективам репликаторов размножаться, потому что из
одной капельки, окруженной мембраной, могут появиться две. Во-вторых, переход
от мира РНК к белковому миру.
Опять-таки, ДНК
в этом смысле неинтересна, потому что это тупая молекула, чисто для
хранения информации. Достоинство ДНК в том, что она более точно воспроизводится,
поэтому как долговременная память она лучше, чем РНК. Мы это можем наблюдать
прямо сейчас, потому что вирус SARS-COV-2, вызывающий ковид, мутирует очень
быстро именно потому, что он РНКовый. Есть теория Кунина, что ДНК изобрели вирусы.
Он через сравнение белков, которые копируют разные типы нуклеиновых кислот, ДНК
и РНК, в разных направлениях, показывает, что функция копирования РНК
более древняя, потом появилась функция делать ДНК из РНК, а потом уже появилась
функция копирования ДНК. И это вещь, которую уже можно пытаться не из
головы сочинять, а посчитать, сравнивая белки и рассматривая возможные
сценарии появления функций в этих семействах белков.
Переход от
нуклеинового мира к белковому — это, по-моему, в каком-то смысле
штука более сложная, чем возникновение жизни (во всяком случае, интеллектуально).
Ну, хорошо, бог с ними, с вероятностными оценками, но там хотя бы
принципиально понятно, как это могло быть, а здесь приходится труднее,
потому что в том синтезе белка, который мы видим, участвует сразу очень
много разных молекул. Ну, рибосомы — ладно, рибосомы — это РНКовый остов
и много белков. Раньше думали, что РНКовый остов нужен как скелет,
а собственно функции удлинения белка, прочитывания очередного триплета
делают белки. Оказалось, что на самом деле основные ферментативные функции
в рибосоме выполняет как раз РНК, которая является тем самым рибозимом.
И это еще одно сильное соображение в пользу первичности РНКового
мира, потому что реакция синтеза белка выполняется РНКовым ферментом. Но там
есть еще куча всяких игроков, которые обеспечивают, собственно, специфичность
этого синтеза, и довольно трудно себе представить (во всяком случае,
мне), как могли бы быть устроены промежуточные полезные шаги.
— Минутку… Там есть некий сложный механизм…
— Механизм синтеза
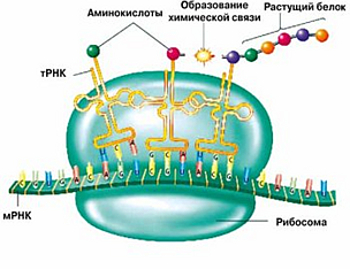
белка несложный. Там просто много участников, сложность в этом. Во-первых,
имеется сама РНКовая матрица (мРНК). В данном случае никакой не рибозим,
а просто матрица, в которой записано, какой белок должен быть
синтезирован. И есть таблица соответствий троек нуклеотидов в мРНК
и аминокислот, которые присоединяются к растущему белку. Это
и есть таблица генетического кода. От нее бывают отклонения, явно более
поздние, но в принципе она почти универсальная. Во-вторых, есть рибосома.
Это довольно сложная конструкция, состоящая из РНКовой центральной части
и белков, которые ее окружают. Рибосома осуществляет реакцию присоединения
аминокислоты, причем не абы как, а в зависимости от того, какой триплет
сейчас читается.
«Википедия»
— Это и есть самый сложный элемент?
— Не-не-не.
Я не сказал, как обеспечивается специфичность, как реализуется таблица
генетического кода. Я ее описал как мысленный конструкт, но не как работающий
механизм.
Итак, дальше…
В-третьих, есть транспортные РНК (тРНК). Это маленькие РНКовые молекулы,
которые подтаскивают аминокислоты к растущему белку. Каждая из них соединена
с одной фиксированной аминокислотой. Таким образом, их должно быть как
минимум 20, но на самом деле бывает больше. Разные тРНК могут присоединять одну
и ту же аминокислоту, но одна тРНК присоединяет только одну аминокислоту.
Это верно на 95%, как всё в биологии. Я могу рассказать, какие бывают
исключения, но это отдельный разговор.
И вот
очередная тройка экспонирована в нужном сайте рибосомы, приплывают разные
тРНК, тыкаются. Если тРНК соединилась с нужным кодоном (путем уотсон-криковского
взаимодействия, просто у нее есть в нужном месте антикодон,
комплементарный кодону), то аминокислота, которая висела на этой тРНК, присоединяется
к растущему белку. Это реакция удлинения белка, которую делает рибосома.
Наивный читатель
подумал бы, что я рассказал механизм реализации генетического кода.
— Наивный главный редактор — тоже…
— А на самом деле
нет, потому что я не сказал, каким образом получается так, что каждая тРНК
имеет свою собственную аминокислоту. На этот случай есть специальные белки,
которые называются аминоацил-тРНК-синтетазы. Эти белки специфичны для каждой
аминокислоты, специфичны для тРНК данного типа, причем сразу для всех, соединенных
с данной аминокислотой. На каждую аминокислоту имеется только одна
аминоацил-тРНК-синтетаза. Она навешивает правильные аминокислоты на правильные
тРНК. И генетический код реализован не в рибосоме, не в тРНК,
а в аминоацил-тРНК-синтетазах. Нетривиальная специфичность возникает
здесь. Всё остальное — чистая механика.
И как эту
хрень собрать из маленьких естественных шагов? Есть люди с богатой фантазией,
которые утверждают, что примерно представляют, как это может быть. Мне немножко
труднее. Но на самом деле понятно. Есть всякие соображения о том, что если
есть рибозим, то хорошо бы у него аминокислота была в качестве
кофактора, потому что рибозим становится более эффективен. Вот у нас уже появилось
взаимодействие РНК с аминокислотами. Потом может оказаться, что еще лучше,
если этот кофактор — не одна аминокислота, а какой-нибудь коротенький
пептид. Потом может оказаться, что какие-то предки тРНК подтаскивали эти аминокислоты,
взаимодействовали с рибозимом и засаживали эту аминокислоту
в нужный пространственный карман. В общем, можно какие-то слова произносить,
но, в отличие от истории с репликаторами, где хотя бы какие-то этапы
реализованы экспериментально, здесь всё остается на уровне рассказов.
Тут
и начинается кунинский мультиверс и вся эта черная магия.
— Выглядит устрашающе для постороннего человека. Как
это всё заработало?
— Я и говорю.
Для меня интеллектуальный вызов не в том, чтобы представить себе, как
жизнь родилась (в нашем определении репликаторов, способных
к дарвиновской эволюции), а в том, как она из РНКовой стала
РНК-белковой. Это прикольно. Есть книжка Михаила Никитина о происхождении
жизни, там всё это гораздо более подробно и внятно толкуется.
— И все-таки у меня складывается ощущение,
что жизнь — вещь редчайшая. Кто-то считает, что жизнь
зарождается на каждой глыбе с водой, летающей в космосе
у подходящей звезды. В это как-то не верится.
— Ну-у-у…
Во-первых, это действительно вопрос веры. Никакого способа узнать у нас
нет. Во-вторых, никто не отменял панспермию. Происходит одно редкое событие,
потом жизнь просто размножается, летая от планеты к планете…
— Панспермия мало что дает, потому что на нее
мало времени. Вселенная-то всего ничего существует. Примерно в три
раза дольше, чем время эволюции на Земле.
— Ну, мы не знаем.
На близких расстояниях панспермия могла работать. Если это случилось достаточно
рано, когда всё было близко…
— Невозможно.
— Астрофизика —
наука еще более магическая, чем биология, я ее обсуждать не готов.
— Там всё просто!
— Просто, только
правды не найдешь.
Предсказание
состоит в следующем: если мы обнаруживаем жизнь где-то относительно
недалеко от Земли, то, скорее всего, это будет жизнь земного типа — ровно
потому, что она не зарождалась независимо. Мне кажется более вероятным, что
у них общий источник.
— Может быть.
— А где-нибудь на
другом конце Вселенной — там может быть всё что угодно.
— Какова роль
случайности? Почему в таблице именно эти нуклеотиды?
— Неизвестно,
естественно. Но есть совершенно чудесные опыты, когда люди меняют таблицу
генетического кода, добавляют новые аминокислоты, которые специфично
встраиваются, добавляют новые нуклеотиды, которые специфично копируются… Современная
синтетическая биология показывает, что конкретный выбор «букв» вроде бы
необязателен. Однако есть некоторые общие принципы. Аминокислоты должны быть
одной хиральности, иначе белки не будут сворачиваться.
Более того,
школьное представление о том, что в биологии есть 20 аминокислот, не
очень точное. В химии аминокислот не 20, а сотни, и биологически
релевантных аминокислот больше, чем 20, они просто в белках оказываются
другим способом, путем модификации тех аминокислот, которые уже вставились.
Более того, вставляются не 20 аминокислот, а больше. Есть селеноцистеин,
который вставляется очень специфично, по другому механизму, в конкретные
белки в конкретных местах. Это расширение генетического кода, но при этом
абсолютно механистическое. Есть небольшое отклонение от генетического кода,
которое позволяет кодировать пиролизин. Это позднее добавление, специфичное для
определенной группы архей, но оно могло бы возникнуть и раньше.
А есть много аминокислот, которые получаются вторичной модификацией тех,
которые уже вставились.
Есть совершенно
чудесное письмо Крика конца 1950-х годов. (Я студентам всегда велю его
читать, но, по-моему, никто этого не делает.) Тогда начали задумываться, что
должен быть генетический код. Еще не было понятно, как он устроен…
— Гамов!
— Гамов сказал,
что на эту тему надо думать, предложил абсолютно дурацкую модель, причем
с самого начала было ясно, что она дурацкая. Но он молодец, он первым сформулировал,
что такая проблема есть.
Так вот, есть
письмо Крика членам РНКового клуба.(Их там было несколько человек, они
обменивались письмами.) Крик обдумывает, какие именно аминокислоты генетическим
кодом должны кодироваться, а какие вторичны. Из каких-то косвенных соображений
он пытается это понять. Химический состав белков был известен. Если вы отрежете
от себя кусок и посмотрите химический состав, то там будет очень много
аминокислоты гидроксипролин, потому что она входит в состав коллагена,
входящего в состав соединительной ткани. Но гидроксипролина нет
в генетическом коде —, это производная пролина, который кодируется. Если
вы наивно будете думать, что кодироваться должны аминокислоты, которых много
в куске мяса, вы приплывете. Дальше у него соображения, что
у животных гидроксипролин есть, а у бактерий его нету вовсе,
поэтому маловероятно, что он есть в генетическом коде. Гидроксипролин есть
только в коллагене, его нет в других белках. Стало быть, это
вторичная вещь. Куча таких рассуждений — и удивительно, что Крик всё
угадал! Это очень поучительный текст. Один из очень немногих примеров
в биологии, когда оказывается полезно просто подумать.