Происхождение жизни. Следующие миллиарды лет
17.05.2022
Источник: Троицкий вариант,17.05.2022, Михаил Никитин; Борис Штерн

Борис Штерн
Как и обещали, мы
продолжаем серию интервью о происхождении жизни. Первое было
с Михаилом Гельфандом, второе — с Александром Марковым.
Теперь главный редактор Борис Штерн беседует с Михаилом Никитиным,
написавшим знаменитую книгу как раз на данную тему.
— Напоминаю, точка, с которой начались предыдущие интервью, —
возникновение репликатора — некой конструкции, которой, скорее всего, была
молекула РНК, умеющая воспроизводить себя, может быть, с небольшими
дефектами, что дает наследственность и изменчивость. И с этого
стартует дарвиновская эволюция. Михаил, теперь изложите ваш взгляд на проблему
первого репликатора. Есть точка зрения, что вероятность его возникновения
чудовищно мала.

Михаил Никитин
— Да. Первый репликатор,
зачем он нужен? Мы знаем единственный механизм, который может порождать более
сложные системы из более простых — это дарвиновская эволюция, работающая путем
мутаций и естественного отбора. Чтобы дарвиновская эволюция шла, нужны
некие системы, способные размножаться — порождать свои собственные копии, не
идеальные. Среди этих копий будет дальше идти отбор по скорейшему размножению.
Простейшая единица, представленная в современном мире, способная размножаться, —
это прокариотная клетка бактериального или архейного типа. Но такая
клетка — это довольно сложная система, она должна содержать как минимум
около полутора тысяч белков, белки должны быть собраны в супрамолекулярные
комплексы, такие как рибосома и роторная АТФаза, нужен геном из более
миллиона нуклеотидов, который будет всё это кодировать, нужна мембрана, которая
будет окружать клетку. То есть клетка — это заведомо хороший репликатор,
но она довольно сложная, и получить клетку без дарвиновской эволюции путем
случайной самосборки очень-очень маловероятно. Так считал еще Фред Хойл
в 1970-е годы, и получается, что размеров видимой Вселенной
и времени ее существования для этого принципиально недостаточно.
Более простые
самореплицирующиеся единицы тоже известны — это вирусы. Но все вирусы являются
паразитами клеток, они несамостоятельны. И теория мира РНК в конце
1970-х — в 1980-е годы попыталась это как-то примирить: нащупать
существование репликаторов, по сложности сравнимых с вирусами, но при этом
не нуждающихся в клетках. Ключевая гипотеза о мире РНК состоит
в том, что они не содержат белков и ДНК, не используют кодируемый
белковый синтез, и из трех типов полимеров — ДНК, РНК, белки —
в них используются только РНК. Потому, что только РНК может эффективно
выполнять функцию катализа, т. е. работать ферментом — то, что сейчас
делают белки, — и хранить наследственную информацию, с чем ДНК
справляется лучше, но РНК, тем не менее, служит наследственным материалом
многих вирусов до сих пор.
Основная форма первичного
репликатора в этой концепции — молекула РНК, способная копировать саму
себя. То есть РНК-фермент рибозим с активностью РНК-зависимой
РНК-полимеразы. Для его существования нужна экологическая ниша, где будут
доступны готовые нуклеотиды для репликации — это тоже вопрос, который надо
решать. С 1990-х годов такой репликатор пытались получить
экспериментально, отбором РНК с нужными каталитическими свойствами. Успехи
за двадцать с лишним лет довольно скромные. Молекулы РНК, которые
сколько-нибудь эффективно копируют другую РНК, — они довольно крупные, больше
200 нуклеотидов, и получить их случайной самосборкой тоже не очень просто
(хотя, конечно, проще, чем клетку). Но, главное, они недостаточно точны
и эффективны, чтобы сделать собственную копию даже в идеальных условиях.
Поэтому неферментативная
репликация была придумана как способ обойти эти проблемы
с рибозимами — РНК-полимеразами. Например, в каких-то условиях
на минеральной подложке новая РНК может собираться не только случайно, но
и на матрице уже существующей цепи РНК. Для катализа этой реакции
в простейшем случае нужен только магний, а рибозимов и тем более
белковых РНК-полимераз не нужно. В экспериментах неферментативная
репликация дает совсем коротенькие молекулы РНК — 10–15, максимум 20
нуклеотидов — и довольно сильно страдает от неточностей. Но она
как-то работает, и можно предположить, что неферментативная репликация предшествовала
первым рибозимам. Она позволяет начать дарвиновскую эволюцию еще раньше, чем
в классическом мире РНК.
У меня есть одна идея,
которую пока химики, работающие с рибозимами, экспериментально не
проверяли, насчет того, каким мог быть первый рибозим. Для неферментативной
репликации первый рибозим, который мог быть поддержан естественным отбором, —
это не полимераза. Это рибозим с противоположной активностью —
экзонуклеаза, отрезающий отдельные звенья от концов цепочки РНК. Как рибозим,
разрушающий цепочки, может способствовать их копированию? Да очень просто:
в любой современной системе копирования нуклеиновых кислот есть ферменты
с такой активностью. Они называются «проверочные экзонуклеазы», отрезающие
с конца растущей цепочки не все нуклеотиды подряд, а только
ошибочные, не комплементарные нуклеотиду матричной цепи. А основные
проблемы с неферментативной репликацией РНК — это как раз низкая
точность, которая эффективно компенсируется такой проверочной экзонуклеазой,
и низкая скорость. А низкая скорость на 90% следует из низкой
точности: к неправильно присоединенному нуклеотиду очень долго сложно
и маловероятно присоединить следующий правильный. То есть на каждом месте
ошибки неферментативная репликация буксует. Рибозим — проверочная
экзонуклеаза, наверное, может быть гораздо проще, чем рибозим-полимераза, — не
200 нуклеотидов, а меньше 100, даже порядка 50, и его появление
гораздо вероятней. Эта идея пока экспериментально не проверена, но мне она
кажется красивой, кажется, что это способ получить дарвиновскую эволюцию как
можно раньше.
— Есть ли какой-то шанс проверить это экспериментально?
— Конечно, такие работы идут
с 1990-х годов — по получению рибозимов путем искусственного отбора из
большой библиотеки молекул РНК со случайными последовательностями. Так было
получено много вариантов рибозимов-полимераз, рибозимов-лигаз и рибозимов
с разными другими ферментативными активностями, вплоть до какой-нибудь
фотолиазы и алкогольдегидрогеназы.
От самокопирования РНК
к синтезу белков
— Теперь я предлагаю пройтись по узким местам. Мы обсудили первое
узкое место — возникновение репликатора. Следующее — Михаил Гельфанд отнес
к узким местам синтез белка.
— Появление рибосомы
и кодирование белка.
— С другой стороны, Александр Марков объяснил, что там в рибосоме
тоже просматривается ядро из РНК, которое могло этот переход облегчить.
— Да, Александр уже рассказал.
То, что я могу сказать, что древнее ядро большой субъединицы рибосомной
РНК — пептидил-трансферазный центр — в ранние эпохи могло работать
отдельно от малой субъединицы, содержащей декодирующий центр. То есть не
пользоваться матричной РНК и не иметь такого мощного средства контроля за
последовательностью создаваемого белка. Но! Это всё равно могло быть поддержано
естественным отбором, если аминокислотный алфавит был гораздо меньше —
в пределах от двух до четырех аминокислот. Поддержан тремя основными
способами. Первый способ: производство гомополимерных пептидов, т. е.
повторять одну аминокислоту всё время. Способ второй — чередовать аминокислоты
— две, три или четыре — какими-то простейшими способами, типа АБ-АБ-АБ. Способ
третий — случайно комбинировать две, три или четыре аминокислоты. При таком
маленьком аминокислотном алфавите белки, а точнее, пептиды, поскольку они
не смогут компактно и однозначно свернуться, будут иметь воспроизводимые
физико-химические свойства, даже если комбинировать две-три аминокислоты
случайно. И очевидный класс пептидов, который может быть поддержан
естественным отбором, который может быть полезен миру РНК и синтезируется
таким образом, — это катионные пептиды, пептиды с положительным зарядом,
содержащие аминокислоты типа лизина и аргинина. Они притягиваются
к отрицательно заряженным РНК (все РНК отрицательно заряжены), помогают
компактному устойчивому сворачиванию рибозимов и повышают активность
рибозимов почти любых типов. Это всё экспериментально проверено. Вот такие
вспомогательные катионные пептиды могли быть продуктом маленькой простенькой проторибосомы,
не имеющей малой субъединицы, не имеющей матричной РНК, не имеющей
декодирующего центра, и имеющей какую-то маленькую часть от современной
большой субъединицы рибосомной РНК.
В экспериментах большую
субъединицу рибосомной РНК кишечной палочки удалось сократить примерно
с 3 тыс. нуклеотидов до 600, т. е. в пять раз,
и показать, что такой огрызок, содержащий пептидил-трансферазный центр
и ничего больше, тем не менее, связывает транспортную РНК, несущую
аминокислоту, и проводит пептидилтрансферазную реакцию. То есть пептиды
соединяет. И эта простая проторибосома дальше могла пошагово, когда каждый
шаг поддержан естественным отбором, усложняться, расширять аминокислотный
алфавит, переходить от случайного чередования аминокислот к закономерному
и, в конце концов, на стадии, по-видимому, 6-аминокислотного алфавита
перейти к кодируемому белковому синтезу. И в структуре
рибосомных белков, и в структуре транспортных РНК, присоединяющих
к ним аминокислоты, можно довольно уверенно заключить, что такие
аминокислоты, как глицин, аланин и пролин, точно были на стадии
некодируемого пептидного синтеза. Скорее всего, на этой же стадии были аспартат,
валин и какие-то положительно заряженные аминокислоты — либо аргинин, либо
какие-то его более простые аналоги, не существующие в современных белках.
То есть минимальный аминокислотный алфавит для того, чтобы получить белки, способные
самостоятельно свернуться, — это глицин, аланин, пролин, аспартат, валин
и одна положительно заряженная аминокислота.
— Понятно, то есть с этого синтез белков стартовал, потом усложнился,
и добавились остальные аминокислоты.
— Да
— Видимо, это самое начало эволюции, которое заняло какой-то неразрешимо
короткий отрезок времени.
— Да, это прошло
в пределах первых миллионов лет.
Прокариоты заселяют
и преобразуют мир
— Дальше прошло около двух миллиардов лет до появления первой эукариотной
клетки. В эти два миллиарда лет что-нибудь еще радикальное произошло?
— За два миллиарда лет от
первой прокариотной клетки до первой эукариотной клетки, конечно, произошли
некоторые интересные события. Эволюция шла не так быстро, как в начале, до
появления самых первых клеток, но прокариоты наращивали разнообразие, их
становилось всё больше и больше, они освоили всю планету. Если первая
клетка была обитателем каких-то редких геотермальных водоемов типа грязевых
котлов и у нее была очень узкая экологическая ниша, то ее потомки
освоили все моря — поверхность и глубины, — пресные воды и проникли
в толщу земной коры. В современной земной коре бурение показывает,
что она заселена бактериями до глубин 4–5, а иногда даже 6 км, причем
распространение жизни в глубину ограничено только ростом температуры — на
глубине 6 км температура уже около 80 °C. Бактерии и археи, заселяющие
толщу земной коры, имеют чудовищную биомассу, превосходящую массу всей жизни на
поверхности, включая деревья. Глубинная биосфера живет медленно
и малопродуктивно, но биомасса ее огромна.
За два миллиарда лет до
появления эукариот жизнь распространилась из луж по всей толще морей
и земной коры, нарастила разнообразие и освоила некоторые новые
биохимические процессы, прежде всего кислородный фотосинтез, который радикально
преобразил планету. Это именно тот процесс, который делает обитаемую планету
наблюдаемой на космических расстояниях. Сейчас приступает к работе
космический телескоп «Джеймс Уэбб», и в его научные задачи входит
спектральный анализ атмосфер транзитных экзопланет на расстояниях до сотни
световых лет, насколько я помню, и, таким образом, он сможет отличить
живую планету, с атмосферой, похожей на земную, обогащенную кислородом
из-за фотосинтеза, от безжизненных планет с преобладанием азота
и углекислого газа в атмосфере. Два не очень стабильных на
геологических временах газа, выделяемых живыми организмами, — метан
и кислород. Метан преимущественно накапливается в атмосфере до
появления на планете кислородного фотосинтеза, после чего накапливается уже
кислород. И то и другое может быть обнаружено современными
телескопами на расстояниях до 100 световых лет.
Видимо, кислородный фотосинтез
сыграл важную роль в появлении эукариот, которые по своей биохимии
довольно однообразны по сравнению с прокариотами. Но у эукариот есть
один биохимический путь, которого бактерии не придумали. Это синтез стеролов,
прежде всего холестирола. Стеролы входят в состав эукариотных клеточных
мембран — придают им повышенную гибкость и текучесть и помогают таким
эукариотным процессам, как фагоцитоз и ошнуровка и слияние мембранных
пузырьков. Фагоцитоз — это поглощение эукариотной клеткой какой-то твердой
пищевой частицы, которая окружается мембраной и проваливается внутрь
клетки. Он позволяет переваривать эту твердую пищу в каком-то
контролируемом объеме. Прокариоты питаются жидкой пищей, растворимыми
молекулами. Только эукариоты смогли питаться твердыми частицами, прежде всего целыми
прокариотными клетками. То есть в прокариотном мире первые эукариоты были
сверххищниками, которые могли съесть кого угодно.
— Вернемся немного назад. Насколько неизбежным было появление
кислородного фотосинтеза, который преобразовал планету? Раз уж возникла
прокариотная жизнь, насколько вероятным будет появление кислородной атмосферы?
— Я могу предположить, что это
достаточно неизбежно. Это следствие нескольких последовательных ресурсных
кризисов. Среди прокариот существует несколько вариантов бескислородного
фотосинтеза. Вообще, весь фотосинтез — это использование энергии света для
восстановления углекислого газа и производства из него органических
молекул, используя электроны, отбираемые для него у каких-то других
внешних молекул. В кислородном фотосинтезе этой внешней молекулой является
вода, которая окисляется до молекулярного кислорода. В основных вариантах
бескислородного фотосинтеза источником электронов являются ионы железа Fe2+,
окисляемые до Fe3+, и соединения серы, такие, как сульфиды или сама
молекулярная сера. Сульфиды окисляются до серы, сера окисляется до сульфатов.
Есть варианты бескислородного фотосинтеза, использующие молекулярный водород,
есть варианты с использованием некоторых органических соединений, таких,
как янтарная или щавелевая кислоты. Понятно, что вода доступна в гораздо
большем количестве, чем вышеперечисленное. И по геологическим данным, на
протяжении архея моря были заселены в основном микробами, осуществляющими
железный фотосинтез. Существует много отложений — полосатые железистые
кварциты, они же джеспилиты, они же banded iron formations — это, судя по
всему, следы деятельности железных фотосинтетиков, окислявших двухвалентное
железо, растворенное в морской воде в ту эпоху, и осаждавших его
в виде магнетита и гематита. Причем осаждавших днем и не
осаждавших ночью — поэтому отложения полосатые, состоящие из субмиллиметровых
полосочек.
Но поступление железа
в морскую воду связано с геологической активностью планеты —
с вулканами, с выветриванием базальтов на суше, с деятельностью
черных курильщиков и т. д. А геологическая активность со
временем затухает. Продуктивность биосферы же, с другой стороны, росла по
мере того, как микробы осваивали новые места обитания и распространялись
в том числе по морю. То есть выпадение железа из моря со временем
усиливалось, а поступление железа в море со временем слабело. Рано
или поздно два этих процесса приходят к такому результату, что железо
в море кончается, и микробы, использующие железный фотосинтез, больше
не могут этим заниматься — они влетают в ресурсный кризис.
Кислородный фотосинтез
позволяет использовать неисчерпаемый в масштабах Земли ресурс. Но он требует
гораздо более сложных ферментативных систем, он гораздо более опасный. Если
кислородным фотосинтезом неаккуратно управлять, то продуктом становится не
только кислород, но и такие ядовитые вещества, как перекись водорода
и совсем ядовитые гидроксильные радикалы, которые легко могут убить
клетку, занимающуюся кислородным фотосинтезом. И если посмотреть на то,
как устроен кислородный фотосинтез в клетках, например, цианобактерий — он
обвязан множеством защитных систем, которые по логике работы напоминают защитные
системы на атомных электростанциях: в любой непонятной ситуации глушите
реактор. То есть в цианобактериях есть система аварийного сброса
мембранного потенциала, если есть проблемы с тем, куда мембранный потенциал
дальше тратить. Есть специальные белки хлорофилл-содержащие, которые
активируются при повышении яркости света и которые рассеивают энергию
поглощенного света в тепло, не пуская ее на окисление воды. Есть множество
белков, которые связывают хлорофилл, выпадающий из поврежденных фотосистем, не
позволяя ему проводить какие-то опасные фотохимические реакции. Есть много
протеинкиназ, которые совершают тонкую регуляцию фотосистем, подстраивая их
свойства к интенсивности света потому, что фотосистемы должны эффективно использовать
тусклый свет на рассвете или на закате и в то же время безопасно для
клетки работать днем в ясную погоду, когда интенсивность света возрастает
в тысячи раз.
У цианобактерий всё это
есть. У микробов, занимающихся бескислородным фотосинтезом, железным
и серным, этого почти нет. Кислородный фотосинтез гораздо опасней
и гораздо сложней. И ключевой ко-фактор кислородного фотосинтеза —
это марганцевый кластер — наночастица оксида марганца, содержащая четыре иона
марганца, соединенных с молекулой фотосистемы II, именно ионы марганца
являются катализаторами окисления воды. Если посмотреть на процесс сборки этого
марганец-кислородного кластера, то очень похоже, что он возник как отход
другого типа фотосинтеза, марганец-окисляющего. В архейской морской воде
помимо ионов двухвалентного железа было также заметное количество
двухвалентного марганца. Марганец — более слабый восстановитель, чем железо,
поэтому пока железа было много, можно было жить за его счет, и марганцевым
фотосинтезом никто заниматься не пытался. Когда железо кончилось, следующий
доступный ресурс — это марганец. Ионы Mn2+, окисляющиеся фотосистемами до четырехвалентного
оксида MnO2, тоже могут поддерживать фотосинтез.
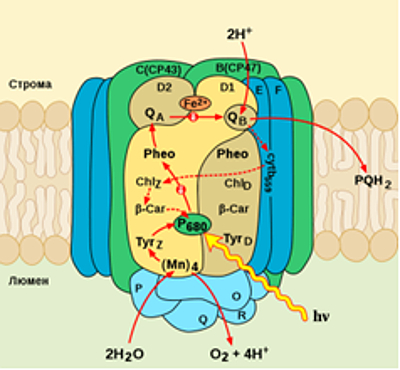
Схема фотосистемы II.
(Mn)4 — Mn-содержащий кластер; Tyrz — тирозин-161 белка D1;
Tyrd — тирозин-161 белка D2; P680 (П680) — специальная пара;
cytb559 — цитохром b559; Chlz, Chld — сопровождающие хлорофиллы а;
β-Car — β-каротин; Qa, Qb — пластохиноны. Пунктиром обозначен циклический
поток электронов
И дальше оказалось, что
если этот оксид марганца прилипает к фотосистеме, то он из отхода
становится средством производства, средством нового типа фотосинтеза, водоокисляющего.
И вот тут у меня есть два интересных соображения.
Во-первых, если посмотреть на
родословное дерево цианобактерий, единственных микробов, способных
к кислородному фотосинтезу, то все нижние ветви на этом дереве —
пресноводные. Освоение цианобактериями морей происходило сильно позже появления
кислородного фотосинтеза и несколькими ветвями независимо. Кислородный
фотосинтез распространился в морях на границе архея и протерозоя,
примерно 2,5 млрд лет назад, но в пресных водах цианобактерии освоили
его гораздо раньше. Глобального эффекта на планете это не имело, потому что
озера и реки гораздо меньше, чем мировой океан, но процесс уже был.
Во-вторых, в пресных
водах с дефицитом железа и соединений серы они могли столкнуться
гораздо раньше, чем в море, — из-за малого объема водоемов. И третье.
Как же цианобактерии преодолели проблему опасности кислородного фотосинтеза?
С химической точки зрения невозможно безопасно перейти от бескислородного
фотосинтеза к кислородному, и вот почему: для окисления железа от каждого
атома железа достаточно отрывать по одному электрону. Фотосистема проводит
через себя по одному электрону на каждый поглощенный квант света. Если мы
отрываем один электрон от молекулы воды, то получаем протон
и гидроксильный радикал — крайне опасное соединение. Если мы от двух
молекул воды отрываем два электрона одновременно, то получаем перекись водорода
— тоже не сахар. Чтобы окислять воду безопасно, надо отрывать от нее сразу
четыре электрона, а фотосистема умеет пропускать их только по одному.
И марганцевый кластер работает как раз промежуточным конденсатором:
сначала фотосистема отделяет от него по одному четыре электрона, и дальше
он в одну стадию возвращает четыре электрона от двух молекул воды, окисляя
их до кислорода. Любые поломки марганец-кислородного кластера, которые можно сделать
в эксперименте с выделенными фотосистемами или внося мутации
в гены фотосистем, приводят к тому, что кислородный фотосинтез
становится опасным прежде всего для самой клетки.
Как эволюция могла поддержать
такое? Безопасного маршрута от бескислородного фотосинтеза к кислородному
нет. Но мне кажется, что безопасный маршрут и не требуется, если первая
функция кислородного фотосинтеза была не такой, как сейчас. Я уже упоминал
некоторые параллели между кислородным фотосинтезом и атомной энергетикой.
А вы помните, для чего строились самые первые атомные реакторы? Вовсе не
для выработки электричества.
— Для наработки плутония.
— Да, для наработки оружейного
плутония. И только когда на деньги военных, которые денег не считали
и не очень заботились о безопасности, реакторная технология
достаточно созрела, появились атомные электростанции. Мне кажется, что подобная
история могла быть и с кислородным фотосинтезом. Что производство
ядовитых активных форм кислорода было не проблемой, а целью первых версий
кислородного фотосинтеза. Это было оружием одних клеток против других — травить
активными формами кислорода каких-то соседей. Химическая война между микробами
в одном сообществе идет и сегодня. В ней используются как
сложные пептидные антибиотики, так и простенькие молекулы типа оксида
азота. И активные формы кислорода в древние эпохи вполне могли
в этом участвовать. То есть какие-то предки цианобактерий жили
в пресных водах, снабжались энергией с помощью бескислородного
фотосинтеза, но при этом у них была какая-то предковая фотосистема II,
содержащая не четыре атома марганца, а один или два, которая могла
производить активные формы кислорода для подавления соседей других видов. То
есть кислородный фотосинтез мог сначала возникнуть как оружие. И только
после его совершенствования и совершенствования средств защиты от активных
форм кислорода он мог стать основой энергетики клетки и процессом, преображающим
планету.
— С этим кислородным фотосинтезом получается интересная, прямо-таки
детективная история. Она выглядит довольно сложной, и странно, как
эволюция умудрилась на это наткнуться. Правильно ли я понимаю, что жизнь
все-таки должна была наткнуться на кислородный фотосинтез из-за его огромной
выгоды? Особенно в связи с исчерпанием доступных железа
и марганца.
— Смотрите. Возникновение
кислородного фотосинтеза и его глобальное распространение — это два разных
события, которые разделены по времени миллиардом или половиной миллиарда лет.
Бактерии с кислородным фотосинтезом всю вторую половину архея были
ограничены пресными водами. В отложениях позднего архея иногда находят
следы кратковременного присутствия кислорода, которые у биологов получили
название «кислородные дуновения» (oxygen whiffs), связанные как раз
с континентальными пресноводными отложениями. То есть кто-то немного
выделял кислород в озерах, но в морях они не были
конкурентоспособными по сравнению с местными железоокисляющими
фотосинтезирующими микробами. И только когда железо в море кончилось,
спустившиеся из озер и рек цианобактерии смогли освоить моря, вытеснив
железоокисляющих фотосинтетиков. Последние сейчас живут во всяких экзотических
местах — в околовулканических озерах, где мало кислорода, но много железа
из земной коры. А когда-то они занимали весь Мировой океан.
Эукариоты: еще один прыжок
через пропасть
Какое отношение всё это имеет
к эукариотам? Мы немного забросили историю про стеролы, их важность для
эукариотной клетки. Так вот, биосинтез стеролов происходит при участии
молекулярного кислорода. Его надо не так много, как для кислородного дыхания,
но немного надо. Тех малых количеств кислорода, которые есть, например,
в кишечниках животных, хватает кишечным паразитам для синтеза стерола, но
не для дыхания. Но в бескислородном мире стерол синтезировать невозможно.
У всех эукариот стерол исходно есть. Значит, они возникли, когда кислород
уже был. В бескислородном мире они, видимо, не могли бы возникнуть.
И с происхождением эукариот связана огромная загадка: эукариотная клетка,
будучи гораздо сложнее прокариотной, при этом имеет гораздо более крупный,
замусоренный и неупорядоченный геном, очень сложно, дорого и медленно
управляемый. То есть переход от прокариот к эукариотам очень трудно
объяснить просто естественным отбором — в отличие от всех предыдущих
событий, поскольку от возникновения первого репликатора до возникновения первой
бактериальной клетки и до кислородного фотосинтеза всё было поддержано естественным
отбором.
Белки выгоднее, чем рибозимы,
как катализаторы, ДНК — более надежный носитель генетической информации, более
плотные клеточные мембраны позволяют выйти из геотермальных водоемов
в моря и озера, кислородный фотосинтез поддержан естественным
отбором, когда кончилось железо. А в случае с эукариотной
клеткой отличие от прокариотной настолько велико, что поддержать все шаги через
эту пропасть по маленьким камешкам естественным отбором, похоже, нельзя. По
крайней мере, ученые не могут придумать такого сценария.
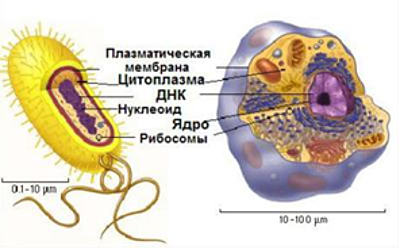
Схематическое сравнение
прокариотной и эукарионтой клеток
Эукариотная клетка возникла
путем симбиоза между археей и бактерией, альфа-протеобактерией, очень
вероятно, что в этом симбиозе поучаствовали еще и гигантские вирусы,
возможно, даже не один. То есть эукариотная клетка представляет собой,
в сравнении с бактериями, огромную, сложную, неуклюжую, очень дорогую
и неэффективную химеру. Тем не менее эукариоты появились. Появились
единственный раз. Многие другие эволюционные процессы мы можем наблюдать
повторно, например многоклеточность в растительном мире возникала больше
десятка раз, многоклеточность грибов возникала три раза, в эволюции
животных тоже довольно много повторяющихся событий, скажем, акула, дельфин
и ихтиозавр похожи друг на друга снаружи, хотя произошли от совершенно
разных предков. А происхождение эукариот — это событие уникальное,
насколько мы можем судить. Ничего подобного на Земле больше не происходило,
а если и происходило, то следы этого стерты настолько начисто, что мы
не видим даже малейших намеков.
Мне лично происхождение
эукариот кажется самым маловероятным шагом в эволюции жизни на Земле. Мне
кажется, что в Галактике могут быть тысячи и миллионы планет,
населенных бактериями, и при этом наша Земля может быть единственной
планетой эукариот. Я, конечно, готов спорить об этом на бутылку коньяка.
К сожалению, пока мы не полетим к другим звездам, решить этот спор
будет трудно. Но мне кажется так.
— Хорошо! Это очень важное заявление, в свете которого вся эукариотная
жизнь — необыкновенная ценность.
— Да.
Животный мир. Цивилизация
как репродуктивная система
— Но дальше, смотрите. Возникли эукариотные клетки. После этого прошло
еще больше миллиарда лет, пока не возникли сложные организмы. То есть дальше
ждали еще какие-то проблемы?
— Да.
— То есть эукариотам нужно было много времени, чтобы научиться
объединяться в сложные организмы. Какие там еще проблемы по дороге?
— Вы, конечно, имеете
в виду под сложными организмами животных? Не растения.
— Да, конечно.
— Многоклеточность растений
возникала несколько раз, а многоклеточность животных всего один раз.
Насколько мы можем судить, многоклеточность животных возникла в позднем
протерозое в эпоху криогения, в эпоху глобальных оледенений, когда
Земля замерзала почти до экватора.
— Сколько это времени назад произошло?
— Примерно 700–750 млн лет
назад, может быть 800.
— То есть это больше 200 млн лет до кембрия?
— Да, задолго до кембрия.
Расхождение разных групп животных, таких, как губки и кишечнополостные,
датированное с помощью молекулярных часов, началось как раз
в криогении. То есть многоклеточность животных возникла тогда. И еще
криогений — период очень быстрого роста содержания кислорода в атмосфере.
Если на границе архея и протерозоя кислород в атмосфере возник вообще
в уловимых количествах, а на протяжении большей части протерозоя его
было довольно мало — несколько процентов от современного количества, — то
в течение криогения его содержание стало расти очень быстро
и к кембрию выросло до половины от современного.
Мне кажется, что тут, как
и с появлением кислородного фотосинтеза, дело во многом
в железе. Появлению кислородного фотосинтеза препятствовало то, что железо
было доступно в морской воде. На границе архея и протерозоя запасы
железа в морской воде были исчерпаны, но оставалось много железа
и других окисляемых минералов на суше, и выделяемый фотосинтетиками
кислород реагировал прежде всего с ними. В протерозое начинается
образование железных руд на суше (так называемые красноцветы).
В протерозое начинается массовое окисление сульфидных минералов на суше,
и получаемая при этом серная кислота течет в моря, где
в огромных количествах появляются сульфаты, начиная с протерозоя.
В архейской морской воде сульфатов практически не было. И этот
восстановительный буфер — сульфиды разных металлов плюс двухвалентное железо
в составе базальтов — очень долго сдерживал рост концентрации кислорода
в атмосфере. Но в течение протерозоя кислородные фотосинтетики росли
и развивались, появлялись более эффективные многоклеточные эукариотные
водоросли, которые плохо поддавались разложению, происходило захоронение неокисленного
органического углерода в морских осадках, что способствовало необратимому
выделению кислорода в атмосферу. Когда фотосинтетики переломили тенденцию
расхода кислорода на окисление минералов на суше, и когда затухающая
геологическая активность замедлила поступление базальтов и сульфидов на
сушу, содержание кислорода в атмосфере начало быстро расти. Открылась
возможность появления многоклеточных животных.
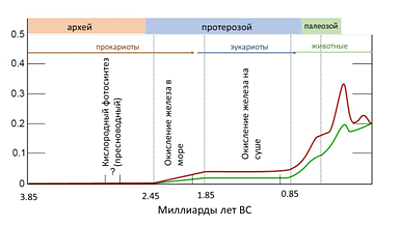
Геологические эры и содержание
кислорода в атмосфере Земли. Красная кривая – максимальные, зеленая –
минимальные оценки. Данные из работы D. Stolper, M. Bender, G.
Dreyfus, Y. Yan, J. Higgins. Science, 23 Sep 2016. Vol 353, Issue 6306
Как и с эукариотами,
у животных тоже есть один общий для всех животных метаболический путь,
использующий молекулярный кислород. И это не дыхание. Это окисление одной
из аминокислот — пролина в гидроксипролин. Это аминокислота, которой нет
в генетическом коде, тем не менее в суммарном составе белков человека
или мыши ее больше, чем таких классических аминокислот, как триптофан
и метионин. Гидроксипролин нужен для синтеза коллагена — ключевого белка
внеклеточного матрикса животных, к которому прикрепляются клетки. Это
базальные мембраны, связки, сухожилия, кости, хрящи, рыхлая соединительная
ткань, нижний слой кожи и т. д. То есть коллагены в организме
животных встречаются везде. И структура коллагенов с его прочными
длинными волокнами (вы знаете, что сухожилия настолько прочны, что
в древнем мире их использовали как тетиву луков?) требует гидроксипролина,
который в свою очередь требует молекулярного кислорода и аскорбиновой
кислоты. Поэтому при недостатке аскорбиновой кислоты этот процесс нарушается —
болезнь называется «цинга», при ней в первую очередь выпадают зубы
и страдают все остальные коллагеносодержащие ткани.
— Возникает впечатление, что путь эволюции до животных, не говоря
о разуме, извилист и тернист. Удивительно, что хватило четырех миллиардов
лет. Это невероятное везение?
— Есть известное уравнение
Дрейка, которое описывает вероятность возникновения разумной жизни
в Галактике. Мне оно не нравится тем, что там есть вероятности, но нет
времени протекания процессов. А от чего зависит время? В случае Земли
очень похоже, что темпы эволюции зависят от доступности железа и скорости
его утилизации фотосинтезирующими организмами. Если бы мы жили на суперземле,
богатой железом и с медленнее затухающей геологической активностью,
то и эволюция от первой клетки до животных, возможно, заняла бы не четыре
миллиарда лет, а все десять или двадцать. Вполне возможно, что на
суперземлях жизнь может когда-нибудь возникнуть. Мы просто успели раньше.
С другой стороны, если бы Солнце было тусклее, фотосинтез шел бы медленнее.
Смотрим на системы красных карликов. Даже при том содержании железа
и затухании геологической активности, которые есть на Земле,
у красного карлика все эти процессы были бы замедлены в несколько раз
из-за меньшей доступности энергии для фотосинтеза. Бескислородные фотосинтетики
десяток миллиардов лет занимались бы осаждением железа, потом еще десяток
миллиардов лет прошел бы до возникновения животных.
— Но там у красных карликов другие проблемы. Зато они живут долго.
Там есть все эти десятки миллиардов лет.
— Но Вселенной меньше
15 млрд лет. Жизнь в системах красных карликов может быть
когда-нибудь возникнет, но мы до этого не доживем.
— В целом впечатление таково, что земная жизнь — нечто совершенно
уникальное, и дело не только в нескольких пропастях, которые пришлось
преодолеть, но и в недостатке времени. На Земле эволюция свершилась
очень быстро и вовремя. Надо иметь в виду, что запас времени невелик:
если бы на Земле не появилась разумная жизнь, то, скорее всего, через миллиард
лет условия для эволюции на Земле исчезли бы. Солнце потихоньку разогревается,
и где-то через миллиард лет Земля превратится в Венеру. Михаил, вам
есть что еще добавить напоследок?
— Да, у меня есть, что
добавить. Вы наверняка слышали про гипотезу Геи (Gaia) Джеймса Лавлока: Земля
как обитаемая планета обладает многими свойствами живого организма, прежде
всего поддержанием гомеостаза. Но на это можно довольно легко возразить, что
Гея не размножается, а живые организмы размножаются. Но если подумать: как
выглядело бы размножение Геи, если бы она вдруг стала это делать? Куда ей
размножаться — здесь она уже заняла всё доступное пространство, все пригодные
для живых организмов места Земли как небесного тела. Для размножения ей нужен
доступ к другим небесным телам. Если мы не берем перенос отдельных
микробов метеоритами, то перенос на другие планеты тысяч видов, включая животных
и растения, возможен только на космическом корабле, построенном
технологической цивилизацией. То есть Гея не размножается только потому, что
она еще маленькая, она еще не достигла зрелости. А мы, человечество, — это
репродуктивная система Геи. Только мы можем помочь ей размножиться
и создать свои копии.
— Ну так я целую книжку об этом написал!
— Да!
— Спасибо за очень информативное интервью, я думаю, не последнее.